«Pierwsze kroki glikolizy
Pierwszym enzymem stosowanym w drugiej fazie glikolizy jest dehydrogenaza 3-fosforan aldehydu glicerynowego; dehydrogenazy to enzymy, które katalizują przeniesienie mocy redukującej z cząsteczki redukującej, która utlenia się do innej cząsteczki, która ulega redukcji (reakcja redoks).Substratami tego enzymu są NAD (dinukleotyd nikotydamidoadeninowy) i FAD (dinukleotyd flawinoadeninowy).
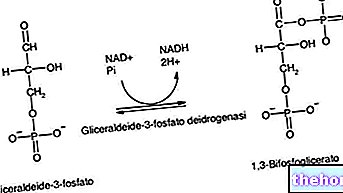
Na tym etapie dehydrogenaza katalizuje konwersję 3-fosforanu aldehydu glicerynowego do 1,3-bisfosfoglicerynianu: w tym samym miejscu katalitycznym grupa aldehydowa jest utleniana do karboksylu, co prowadzi do redukcji NAD+ do NADH, a następnie grupa karboksylowa jest zdolna tworząc wiązanie bezwodnikowe z ortofosforanem. Pierwszy proces jest bardzo egzergiczny (uwalnia energię), podczas gdy drugi jest bardzo egzergiczny (wymaga energii); gdyby nie było miejsca katalitycznego, reakcja globalna nie miałaby miejsca: pierwsza reakcja zachodziłaby z uwolnieniem energii, która byłaby rozproszona jako ciepło, a zatem nie byłaby użyteczna do utworzenia wiązania bezwodnikowego.

Po utworzeniu 1,3-bisfosfoglicerynianu enzym powraca do swojej wyjściowej struktury i jest gotowy do działania na nowym podłożu.

Potem przychodzi kinaza fosfoglicerynianowa co umożliwia przeniesienie fosforylu z 1,3-bisfosfoglicerynianu do cząsteczki ADP; uzyskaliśmy ATP (jeden ATP na każdą cząsteczkę 3-fosforanu aldehydu glicerynowego, a więc dwa ATP na każdą początkową cząsteczkę glukozy), który kompensuje wydatek energetyczny pierwsza faza glikolizy.
Anion arsenianowy (AsO43-) wpływa na szlak glikolityczny, ponieważ może zastąpić fosforan w pierwszej reakcji drugiej fazy glikolizy, dając 1-arsenio-3-fosfoglicerynian, który jest wysoce niestabilny i po uwolnieniu się od katalizatora hydrolizuje, uwalniając „arsenian powracający do obiegu”. Dlatego arsenian naśladuje działanie fosforanu i wchodzi w miejsce katalityczne: w obecności arsenianu nie zachodzi reakcja, w wyniku której powstaje ATP (z 1,3-bisfosfoglicerynianu do 3-fosfoglicerynianu), ponieważ 3-fosforanowy aldehyd glicerynowy przekształca się bezpośrednio w 3-fosfoglicerynian, przy braku dostępnego ATP komórki umierają (zatrucie kwasem arsenowym).
W trzeciej reakcji fazy oksydacyjnej 3-fosfoglicerynian jest przekształcany w 2-fosfoglicerynian w wyniku działania mutaza fosfoglicerynianowa; reakcja obejmuje związek pośredni 2,3-bisfosfoglicerynian.

W następnym kroku interweniuje enzym enolaza który jest w stanie katalizować eliminację cząsteczki wody ze szkieletu węglowego 2-fosfoglicerynianu, otrzymując pirywowany fosfoenol (PEP);

PEP ma wysoki potencjał przenoszenia fosforylu: przenosi się dzięki działaniu enzymu kinaza pirogronianowa, fosforyl do ADP z wytworzeniem ATP, w piątym etapie drugiej fazy, otrzymując pirogronian.

2-fosfoglicerynian i 3-fosfoglicerynian mają niską moc przenoszenia fosforylu, dlatego w celu uzyskania ATP z tych cząsteczek 3-fosfoglicerynian jest przekształcany w 2-fosfoglicerynian podczas glikolizy, ponieważ jest z niej otrzymywany PEP, który jest gatunki o wysokim potencjale przenoszenia.
Zanim przejdziemy dalej, otwórzmy nawias na 2,3-bisfosfoglicerynianie; ta ostatnia występuje we wszystkich komórkach, w których glikoliza zachodzi w bardzo niskim stężeniu (jest produktem pośrednim trzeciej reakcji drugiej fazy glikolizy). Z drugiej strony w erytrocytach 2,3-bisfosfoglicerynian ma stacjonarne stężenie 4-5 mM (stężenie maksymalne), ponieważ mają one dziedzictwo enzymatyczne, które ma za zadanie go wytworzyć; w erytrocytach występuje odchylenie od glikolizy do wytwarzania 2,3-bisfosfoglicerynianu: 1,3-bisfosfoglicerynian jest przekształcany w 2,3-bisfosfoglicerynian w wyniku działania mutaza bisfosfoglicerynianowa (erytrocytów) i 2,3-bisfosfoglicerynian, przez działanie fosfataza bisfosfoglicerynianowa (erytrocyt) staje się 3-fosfoglicerynianem. Następnie w erytrocytach część 1,3-bisfosfoglicerynianu uzyskanego z glikolizy jest przekształcana w 2,3-bisfosfoglicerynian, który następnie powraca na szlak glikolityczny jako 3-fosfoglicerynian; w ten sposób trzeci etap fazy oksydacyjnej glikoliza, z której uzyskuje się ATP.Ilość utraconego ATP jest ceną, jaką erytrocyt jest gotów zapłacić za utrzymanie stężenia 2,3-bisfosfoglicerynianu, którego potrzebują te komórki, ponieważ wpływa on na zdolność „hemoglobiny do wiązania” tlenu.
Widzieliśmy, że w pierwszej reakcji drugiej fazy glikolizy NAD+ jest redukowany do NADH, ale konieczne jest, aby po otrzymaniu pirogronianu NADH uległ ponownej konwersji do NAD+: zachodzi to z fermentacją mlekową (uzyskuje się mleczan) lub przez fermentację alkoholową (w grę wchodzą dekarboksylaza pirogronianowa, która dekarboksyluje pirogronian i dehydrogenaza, która tworzy etanol); fermentacje nie wymagają tlenu (beztlenowce).
Wskutek fermentacji mlekowej kwas mlekowy, jeśli nie zostanie odpowiednio usunięty, gromadzi się w mięśniach i uwalniając H + powoduje mimowolne skurcze mięśni, a tym samym skurcze, mięsień w silnym stresie również może osiągnąć minimalne pH 6,8 .
W cyklu Cori część zmęczenia mięśnia jest przenoszona do wątroby, gdy mięsień jest przeciążony. Załóżmy, że mięsień pracuje bez dopływu tlenu (błędne założenie): jeśli mięsień pracuje umiarkowanie, ATP potrzebne do skurczu jest dostarczane wyłącznie przez glikolizę.Jeśli aktywność mięśnia wzrasta i wymagane jest dodatkowe ATP, przyspiesz metabolizm tlenowy, przekształcając mleczan, który jest w ten sposób usuwany, do glukozy. W rzeczywistości mięsień wykorzystuje metabolizm tlenowy: jeśli jest dostępny tlen, mięsień wykorzystuje przede wszystkim ATP dostarczane przez metabolizm tlenowy, a gdy nie ma już dostępnego tlenu, metabolizm beztlenowy jest przyspieszany przez cykl Cori Cykl ten zakłada, że mleczan jest przenoszony z mięśnia do wątroby, gdzie poprzez wydatkowanie energii powstaje więcej glukozy, która wraca do mięśnia.W trakcie tego cyklu część ATP zużywanego w mięśniu jest dostarczana przez wątrobę, która, poprzez proces glukoneogenezy jest zdolny do wytwarzania glukozy, która może być wykorzystana przez mięsień do uzyskania ATP.
Opisany dotychczas metabolizm glukozy nie obejmuje tlenu, ale tlenowy metabolizm glukozy pozwala na uzyskanie 17-18 razy większych ilości ATP niż uzyskiwane na drodze glikolitycznej, a zatem, gdy komórka ma możliwość wyboru między tlenem a ed beztlenowy, faworyzuje ten pierwszy.
W metabolizmie tlenowym pirogronian przedostaje się do mitochondriów, gdzie ulega przemianom i ostatecznie uzyskuje się dwutlenek węgla i wodę; w ten sposób uzyskuje się 34 cząsteczki ATP na każdą cząsteczkę zdegradowanej glukozy.