Edytowane przez dr Giovanniego Chetta
Białka specjalistyczne
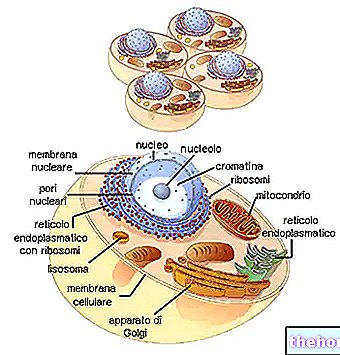
ECM zawiera dużą (i wciąż słabo zdefiniowaną) liczbę wyspecjalizowanych białek niekolagenowych, które zazwyczaj mają wiele domen ze specyficznymi miejscami wiązania dla innych cząsteczek ECM i receptorów na powierzchni komórki. W ten sposób każdy pojedynczy składnik tych białek działa jak „wzmacniacz” kontaktów, zarówno między podobnymi, jak i różnymi cząsteczkami, tworząc nieskończoną sieć biochemiczną zdolną do generowania, modulowania, zmieniania i propagacji, nawet na odległość, milionów i milionów informacje biochemiczne (i energetyczne).
Ważnym wyspecjalizowanym białkiem macierzy zewnątrzkomórkowej jest fibronektyna, glikoproteina o wysokiej masie cząsteczkowej występująca u wszystkich kręgowców.Jest to dimer glikoproteiny złożony z dwóch dużych podjednostek połączonych mostkami dwusiarczkowymi.Fibronektyna wydaje się mieć różny wpływ na wzrost. adhezję komórkową, międzykomórkową oraz z macierzą zewnątrzkomórkową (posiada zdolność wiązania się z komórkami i innymi cząsteczkami macierzy zewnątrzkomórkowej, np. kolagenem, fibryną, heparyną), migracją komórek (komórka może poruszać się do 5 cm dziennie – Albergati, 2004) itp. Najbardziej znana izoforma typu III wiąże się z integryny. Te ostatnie to rodzina białek transbłonowych, które wpływają na adhezję komórek do macierzy zewnątrzkomórkowej poprzez rozpoznanie Sekwencja RGD. Peptydy RGD to łańcuchy aminokwasowe o różnej długości, zaczynające się od prostych tripeptydów, charakteryzujące się sekwencją aminokwasową Arginina-Glicyna-Kwas Asparaginowy. Ta sekwencja aminokwasowa jest wszechobecna w organizmie i bierze udział w wielu funkcjach fizjologicznych.Ta sekwencja RGD reprezentuje jedną ze struktur umożliwiających adhezję poszczególnych linii komórkowych (np. komórek zapalnych) tak, aby mogły pełnić swoją funkcję. Związek między integrynami i RGD indukuje w cytoplazmie szereg reakcji, które obejmują cytoszkielet i inne białka regulujące adhezję, wzrost i migrację komórek.Integryny działają zatem jak mechanoreceptory: przekazują selektywnie i modulowalnie trakcje i napory mechaniczne z ECM do komórki i odwrotnie (Hynes, 2002). Wreszcie, wydaje się prawdą, że włókienka fibronektyny są prawidłowo ułożone i uformowane tylko w obecności napięcia; to rozciąganie jest generowane przez same komórki (Alberts, 2002).
Glukozaminoglikany (GAG) i proteoglikany (PG)
Glukozaminoglikany (GAGS) i proteoglikany (PG) tworzą silnie uwodnioną żelowatą substancję zdefiniowaną w tkankach łącznych, w której mieszczą się i otaczają białka włókniste. Ta forma żelu polisacharydowego jest w stanie z jednej strony pozwolić ECM przeciwstawić się znacznym siłom ściskającym, az drugiej strony umożliwić szybką i stałą dyfuzję składników odżywczych, metabolitów i hormonów między krwią a tkankami.
GAG zwykle wiążą się kowalencyjnie z rdzeniem białkowym, dając początek proteoglikanom (PGS). GAG i PG są zdolne do działania samodzielnie lub w grupach jako receptory dla cząsteczek adhezyjnych lub jako katalizatory procesów biochemicznych na krążących cząsteczkach, takich jak czynniki wzrostu, cytokiny i enzymy krzepnięcia.
Glukozaminoglikany (GAGs) są reprezentowane przez łańcuchy polisacharydowe złożone z jednostek disacharydowych wielokrotnie powtarzanych.Jeden z dwóch cukrów jest zawsze reprezentowany przez aminocukier (n-acetyloglukozamina lub n-acetylogalaktozamina) prawie zawsze siarczan. Drugim cukrem jest zwykle kwas glukuronowy lub jego iduronowy izomer L. Istnieją 4 główne grupy GAG: hialuronany, siarczany chondroityny i siarczany dermatanu, siarczany heparanu, siarczany keratanu.
Łańcuchy polisacharydowe glukozaminoglikanów są wolumetrycznie zbyt sztywne, aby zwijać się wewnątrz zwartych struktur kulistych typowych dla łańcuchów polipeptydowych, a ponadto są wysoce hydrofilowe. zajmują dużą objętość w stosunku do swojej masy i tym samym tworzą znaczne ilości żelu nawet przy niskich stężeniach Duża ilość ładunków ujemnych (GAG to najliczniejsze komórki anionowe, zwykle zasiarczone, wytwarzane przez komórki zwierzęce) przyciąga liczne kationy; wśród nich dominującą rolę odgrywa Na+, który zapewnia pełną pojemność osmotyczną i zatrzymuje ogromną ilość wody w ECM. W ten sposób powstają obrzęki (turgory), które pozwalają ECM przeciwstawić się nawet ważnym siłom ściskającym (dzięki temu np. chrząstka stawu biodrowego może w warunkach fizjologicznych doskonale wytrzymać ciśnienie kilkuset atmosfer).
Wewnątrz tkanki łącznej GAG stanowią mniej niż 10-12% całkowitej masy, jednak dzięki swoim właściwościom wypełniają wiele przestrzeni pozakomórkowych, tworząc pory uwodnionego żelu o różnej wielkości i gęstości ładunków elektrycznych, działając w ten sposób z selektywnych kluczowych punktów lub „serwerów”, przez które regulowany jest ruch cząsteczek i komórek wewnątrz MEC, w oparciu o ich rozmiar, wagę i ładunek elektryczny.
Kwas hialuronowy (hialuronian, hialuronian) jest prawdopodobnie najprostszym z GAG. U człowieka składa się z około 25 000 równych niesiarczanowanych disacharydów i zwykle nie jest połączony z żadnym „rdzeniem białkowym”, dlatego ma nietypową strukturę. Dane eksperymentalne i molekularno-biologiczne potwierdzają, że odgrywa on fundamentalną rolę na poziomie kości i stawów w zakresie odporności na znaczne naciski. Ponadto odgrywa bardzo ważną rolę w wypełnianiu przestrzeni w macierzy zewnątrzkomórkowej podczas rozwoju embrionalnego: tworzy puste przestrzenie między komórkami, do których będą migrować w późniejszych stadiach (Albergati, 2004).
GAG i PG, łącząc się ze sobą, mogą powodować powstanie ogromnych kompleksów polimerowych w ECM. Na przykład cząsteczki aggrecano, które reprezentują większość PG na poziomie stawów, łączą się poprzez wiązania niekowalencyjne z kwasem hialuronowym, dając początek agregatom wielkości bakterii.
Nie wszystkie PG są wydzielane przez ECM, niektóre są integralnymi składnikami błon plazmatycznych. Wśród najbardziej znanych membranowych PG, i syndekanie mają domenę zewnątrzkomórkową złożoną z trzech łańcuchów GAG, podczas gdy uważa się, że domena wewnątrzkomórkowa jest zdolna do reagowania z aktyną cytoszkieletu komórkowego (Alberts, 2002).
Inne artykuły na temat „Fibronektyny, glukozaminoglikanów i proteoglikanów”
- Kolagen i elastyna, włókna kolagenowe w macierzy pozakomórkowej
- Macierz zewnątrzkomórkowa
- Znaczenie macierzy pozakomórkowej w równowadze komórkowej
- Zmiany macierzy zewnątrzkomórkowej i patologie
- Tkanka łączna i macierz zewnątrzkomórkowa
- Powięź głęboka - Tkanka łączna
- Mechanoreceptory powięziowe i miofibroblasty
- Biomechanika powięzi głębokiej
- Postawa i równowaga dynamiczna
- Tensegrity i ruchy śrubowe
- Kończyny dolne i ruch ciała
- Wsparcie zamka i aparat stomatognatyczny
- Przypadki kliniczne, zmiany postawy
- Przypadki kliniczne, postawa
- Ocena postawy - przypadek kliniczny
- Bibliografia - Od macierzy pozakomórkowej do postawy. Czy system łączności jest naszym prawdziwym Deus ex machina?














-cos-esami-e-terapia.jpg)







