Edytowane przez dr Giovanniego Chetta
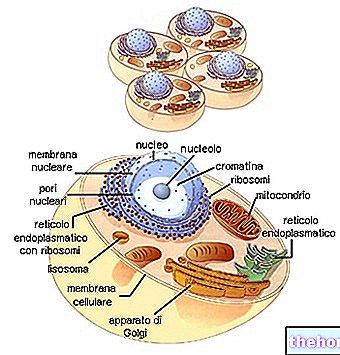
ECM jest ogólnie opisywany jako złożony z kilku dużych klas biomolekuł:
- Białka strukturalne (kolagen i elastyna)
- Białka specjalistyczne (fibrylina, fibronektyna, laminina itp.)
- Proteoglikany (aggrekany, syndekany) i gluzaminoglikany (hialuroniany, siarczany chondroityny, siarczany heparanu itp.)
Białka strukturalne
Kolageny tworzą najbardziej reprezentowaną rodzinę glikoprotein w królestwie zwierząt. Są to najbardziej obecne białka w macierzy zewnątrzkomórkowej (ale nie najważniejsze) i są podstawowymi składnikami właściwej tkanki łącznej (chrząstki, kości, powięzi, ścięgien, więzadeł).
Istnieje co najmniej 16 różnych typów kolagenu, z których typy I, II i III są najbardziej obecne na poziomie typowych włókienek (typ IV tworzy rodzaj siateczki, która stanowi główny składnik blaszek podstawnych).
Kolageny są w większości syntetyzowane przez fibroblasty, ale komórki nabłonkowe są również w stanie je syntetyzować.
Włókna kolagenowe nieustannie oddziałują z ogromną ilością innych cząsteczek macierzy zewnątrzkomórkowej, stanowiąc biologiczne kontinuum fundamentalne dla życia komórki.Związane z nimi kolageny we fibrylach odgrywają dominującą rolę w tworzeniu i utrzymywaniu struktur odpornych na siły napięcia, jest prawie nieelastyczny (glukozaminoglikany pełnią funkcję odporności na ściskanie).W pewien sposób kolagen jest wytwarzany i ponownie metabolizowany w funkcji obciążenia mechanicznego, a jego właściwości lepkosprężyste pociągają za sobą, jak zobaczymy w akapicie „Wiskoelastyczność powięź” , duży wpływ na postawę mężczyzny. Jako kolejny dowód na zdolność kolagenu do zmian pod wpływem czynników środowiskowych, zakładając m.in. różne stopnie sztywności, elastyczności i odporności, istnieją kolageny, określone terminem FACIT (Fibril Associated Collagen with Interrupted Triple helices) zdolne funkcjonować jak proteoglikany (opisane w akapicie „Glukozaminoglikany i proteoglikany”).
Włókna kolagenowe dzięki powłoce PG/GAG (proteoglikany/glukozaminoglikany) posiadają właściwości biosensorów i bioprzewodników: względne ładunki elektryczne skutkują większą zdolnością wiązania wody i wymiany jonów, a tym samym większą pojemnością elektryczną.
Wiemy, że każda siła mechaniczna zdolna do wywołania deformacji strukturalnej napręża wiązania międzycząsteczkowe, wytwarzając niewielki strumień elektryczny, czyli prąd piezoelektryczny (Athenstaedt, 1969). W takich przypadkach włókna kolagenowe rozprowadzają ładunki dodatnie na swojej wypukłej powierzchni, a ujemne na wklęsłej, przekształcając się w półprzewodniki (pozwalają na przepływ elektronów po swojej jednokierunkowej powierzchni). Ponieważ energia piezoelektryczna (jak również energia piroelektryczna generowana przez naprężenia termiczne) jest neutralizowana przez krążące jony w bardzo krótkim czasie (ok. 10-7 - 10-9 sekund), układ PG/GAG na sygnale ma decydujące znaczenie dla propagacji powierzchni sygnału fibryli, pełniąc rolę „repetytorów” impulsu elektrycznego. W szczególności, okresowość podłużna ok. 64 nm (która pod mikroskopem optycznym pojawia się jako smuga) pozwala na prędkość propagacji impulsu równą około 64 m/s (odpowiadającą prędkości przewodzenia szybkich włókien nerwowych) – Rengling, 2001. Silny moment dipolarny włókienek kolagenowych i ich zdolność rezonansowa (właściwość wspólna dla wszystkich struktur peptydowych), a także niska stała dielektryczna MEC, ułatwiają transmisję sygnałów elektromagnetycznych.Dlatego trójwymiarowa i wszechobecna sieć kolagenowa ma również szczególną charakterystykę przewodzenia sygnałów bioelektrycznych w trzy wymiary przestrzeni, oparte na względnym rozmieszczeniu włókienek kolagenowych i komórek, w kierunku aferentnym (od macierzy zewnątrzkomórkowej do komórek) lub odwrotnie, odprowadzającej.
Wszystko to reprezentuje system komunikacji komórkowej MEC w czasie rzeczywistym i takie biosygnały elektromagnetyczne mogą prowadzić do ważnych zmian biochemicznych, na przykład w kości, osteoklasty nie mogą „trawić” piezoelektrycznie naładowanej kości (Oschman, 2000).
Na koniec należy podkreślić, że komórka, co nie jest zaskakujące, wytwarza nieprzerwanie i ze znacznym nakładem energii (ok. 70%) materiał, który koniecznie musi być wydalony, głównie poprzez wyłączne przechowywanie protokolagenu (biologicznego prekursora kolagenu) w określonych pęcherzyki (Albergati, 2004).
Zdecydowana większość tkanek kręgowców wymaga jednoczesnej obecności dwóch istotnych cech: siły i elastyczności. Rzeczywista sieć włókien sprężystych, zlokalizowana wewnątrz macierzy zewnątrzkomórkowej tych tkanek, pozwala na powrót do warunków początkowych po silnych pociągach.Włókna sprężyste są w stanie co najmniej pięciokrotnie zwiększyć rozciągliwość narządu lub jego części. Długie, nieelastyczne włókna kolagenowe są przeplatane między włóknami elastycznymi w celu precyzyjnego ograniczenia „nadmiernych deformacji spowodowanych naciąganiem tkanek. L”elastyna stanowi główny składnik włókien elastycznych. Jest to niezwykle hydrofobowe białko, o długości około 750 aminokwasów, ponieważ kolagen jest bogaty w prolinę i glicynę, ale w przeciwieństwie do kolagenu nie jest glikowany i zawiera wiele reszt hydroksyproliny, a nie hydroksylizynę. Elastyna jawi się jako prawdziwa sieć biochemiczna o nieregularnym, trójwymiarowym kształcie, złożona z włókien i blaszek, które przenikają ECM wszystkich tkanek łącznych.Występuje w szczególnie obfitych ilościach w naczyniach krwionośnych o właściwościach elastycznych (jest białkiem ECM więcej obecne w tętnicach i stanowią ponad 50% całkowitej suchej masy aorty), w więzadłach, płucach i skórze. W skórze właściwej, w przeciwieństwie do kolagenu, gęstość i objętość elastyny z czasem wzrasta, ale stara elastyna na ogół wydaje się spuchnięta, prawie spuchnięta, często z fragmentarycznym wyglądem i zmniejszeniem składnika. (Pasquali Rochetti i wsp., 2004). Głównymi producentami jej prekursora, tropoelastyny, wydzielanej w przestrzeniach pozakomórkowych, są komórki mięśni gładkich i fibroblasty.
Inne artykuły na temat "Kolagen i elastyna, włókna kolagenowe w macierzy pozakomórkowej"
- Macierz zewnątrzkomórkowa
- Fibronektyna, glukozaminoglikany i proteoglikany
- Znaczenie macierzy pozakomórkowej w równowadze komórkowej
- Zmiany macierzy zewnątrzkomórkowej i patologie
- Tkanka łączna i macierz zewnątrzkomórkowa
- Powięź głęboka - Tkanka łączna
- Mechanoreceptory powięziowe i miofibroblasty
- Biomechanika powięzi głębokiej
- Postawa i równowaga dynamiczna
- Tensegrity i ruchy śrubowe
- Kończyny dolne i ruch ciała
- Wsparcie zamka i aparat stomatognatyczny
- Przypadki kliniczne, zmiany postawy
- Przypadki kliniczne, postawa
- Ocena postawy - przypadek kliniczny
- Bibliografia - Od macierzy pozakomórkowej do postawy. Czy system łączności jest naszym prawdziwym Deus ex machina?