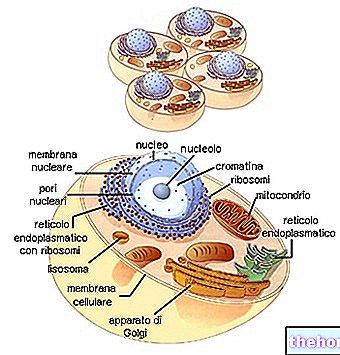
Aby istniała zgodność między informacją polinukleotydu a informacją polipeptydu, istnieje kod: kod genetyczny.
Ogólną charakterystykę kodu genetycznego można wymienić w następujący sposób:
Kod genetyczny składa się z trojaczków i jest pozbawiony wewnętrznej interpunkcji (Crick i Brenner).
Została „odszyfrowana za pomocą systemów translacji otwartych komórek” (Nirenberg i Matthaei, 1961; Nirenberg i Leder, 1964; Korana, 1964).
Jest wysoce zdegenerowany (synonimy).
Organizacja tabeli kodów nie jest przypadkowa.
Trojaczki „bzdury”.
Kod genetyczny jest „standardowy”, ale nie „uniwersalny”.
Patrząc na tabelę kodu genetycznego, należy pamiętać, że odnosi się ona do translacji „RNAm do polipeptydu, dla którego zaangażowane zasady nukleotydowe to A, U, G, C. Biosynteza łańcucha polipeptydowego jest translacją sekwencja nukleotydów w sekwencji aminokwasowej.

Każda trójka zasad RNAm, zwana kodonem, ma pierwszą zasadę w lewej kolumnie, drugą w górnym rzędzie, trzecią w prawej kolumnie.Weźmy na przykład tryptofan (tj. Try) i widzimy, że odpowiedni kodon będzie być w porządku, UGG. W rzeczywistości pierwsza podstawa, U, obejmuje cały rząd pudełek u góry; w tym przypadku G identyfikuje pudełko znajdujące się najbardziej na prawo i czwarty wiersz samego pudełka, w którym znajduje się napis Try. Podobnie, aby zsyntetyzować tetrapeptyd Leucyna-Alanina-Arginina-Serina (symbole Leu-Ala-Arg-Ser) możemy znaleźć w kodzie kodony UUA-AUC-AGA-UCA.

W tym miejscu należy jednak zauważyć, że wszystkie aminokwasy naszego tetrapeptydu są kodowane (w przeciwieństwie do tryptofanu) przez więcej niż jeden kodon. Nie jest przypadkiem, że w przedstawionym przykładzie wybraliśmy wskazane kodony Mogliśmy zakodować ten sam tripeptyd z inną sekwencją RNAm, taką jak CUC-GCC-CGG-UCC.
Początkowo faktowi, że pojedynczy aminokwas odpowiada więcej niż tripletowi, nadano znaczenie losowości, wyrażającej się również w doborze terminu degeneracji kodu, używanego do określenia zjawiska synonimii. Z drugiej strony, niektóre dane sugerują, że dostępność synonimów odnoszących się do różnej stabilności informacji genetycznej wcale nie jest przypadkowa, co wydaje się potwierdzać również odkrycie innej wartości stosunku A + T / G + C na różnych etapach ewolucji. Na przykład u prokariotów, gdzie potrzeba zmienności nie jest zaspokojona przez reguły mendelizmu i neomendelizmu, wzrasta stosunek A + T / G + C. Wynikająca z tego mniejsza stabilność w obliczu mutacji zapewnia większą możliwości zmienności losowej z mutacji genów.
U eukariontów, w szczególności w komórkach wielokomórkowych, w których konieczne jest, aby wszystkie komórki jednego organizmu zachowały to samo dziedzictwo dziedziczne, stosunek A + T / G + C w DNA ma tendencję do zmniejszania się, zmniejszając możliwość mutacji genów somatycznych .
Istnienie kodonów synonimicznych w kodzie genetycznym rodzi wspomniany już problem wielości lub braku antykodonów w RNAt.
Pewne jest, że dla każdego aminokwasu istnieje co najmniej jeden RNAt, ale nie jest równie pewne, czy pojedynczy RNAt może wiązać się z pojedynczym kodonem, czy też może rozpoznawać synonimy obojętnie (zwłaszcza, gdy różnią się one tylko dla trzeciej zasady).
Możemy wywnioskować, że na każdy aminokwas przypada średnio trzy synonimiczne kodony, podczas gdy antykodon to co najmniej jeden i nie więcej niż trzy.
Przypominając, że geny są pomyślane jako pojedyncze odcinki bardzo długich sekwencji polinukleotydowych DNA, jasne jest, że początek i koniec pojedynczego genu muszą koniecznie być zawarte w pamięci.
BIOSYNTEZA BIAŁEK
W różnych częściach DNA następuje otwarcie podwójnego łańcucha i synteza różnych typów RNA.
Podczas etapu ładowania RNAt wiąże się z aminokwasami (wcześniej aktywowanymi przez ATP i specyficzny enzym). Biosyntetyczna „maszyna” nie jest w stanie „skorygować” nieprawidłowo załadowanych tRNA.
RNAr dzieli się następnie na dwie podjednostki i wiążąc się z białkami rybosomalnymi, powoduje powstanie rybosomów.
RNAm, przechodząc przez cytoplazmę, wiąże się z rybosomami, tworząc polisom.Każdy rybosom, przepływając przez przekaźnik, stopniowo zawiera RNAt komplementarne do odpowiednich kodonów, pobierając aminokwasy i wiążąc je z tworzącym się łańcuchem polipeptydowym.
Względnie stabilne RNAt wracają do krążenia. Rybosomy są również ponownie używane, uwalniając już złożony polipeptyd.
Posłaniec, mniej stabilny, ponieważ jest w całości jednosieciowy, jest cięty (przez rybonukleazę) na składowe rybonukleotydy.
W ten sposób cykl trwa, syntetyzując jeden po drugim polipeptydy na informacyjnym RNA dostarczonym przez transkrypcję.